|
|
Post by Dawn2Earth on Nov 25, 2010 22:40:11 GMT -5
This thread is focused towards studies on ancient Kemetian physiognomy i.e. physical appearance.
Variation over time and between regions, affinities to folks elsewhere ancient and modern, etc.
|
|
|
|
Post by Dawn2Earth on Nov 25, 2010 22:42:40 GMT -5
It has been established that (bolded, below):
"The origins of the ancient Egyptian state and its formation have received much attention through analysis of mortuary contexts, skeletal material, and trade. Genetic diversity was analyzed by studying craniometric variation within a series of six time-successive Egyptian populations in order to investigate the evidence for migration over the period of the development of social hierarchy and the Egyptian state. Craniometric variation, based upon 16 measurements, was assessed through principal components analysis, discriminant function analysis, and Mahalanobis D2 matrix computation. Spatial and temporal relationships were assessed by Mantel and Partial Mantel tests. The results indicate overall population continuity over the Predynastic and early Dynastic, and high levels of genetic heterogeneity, thereby suggesting that state formation occurred as a mainly indigenous process."
- Zakrzewski (2007)
So state formation didn't come with any sweeping genetic impact that could be gleaned physically.
Moving on ...
Affinities (or lack thereof) to other peoples hugging the Mediterranean:
"…Finkel (1974, 1978), after a normal equivalent deviate analysis of numerous diachronic ancient “Middle Eastern” cranial series, including some Egyptian, Iranian, Turkish, Syrian, and Greek ones, as well as the Lachish Iron Age series, is able to conclude that this broad region’s populations do not have the characteristics of a single breeding population, i.e., a model viewing this area as a single breeding region with a single morphometric pattern fails. Thus the "Mediterranean race” concept is seen to be questionable-if not invalid-at least on the craniometric level."
- Keita, 1988, An analysis of crania from Tell-Duweir using multiple discriminant functions.
Affinities to the Maghreb
"Epipaleolithic 'mesolithic' Nile Valley remains have these characteristics and diverge notably from their Maghreban and European counterparts in key cranio-facial characteristics (see comments in Keita 1990) although late Natufian hunters and early Anatolian farmers (Angel 1972) shared some of these traits, suggesting late Paleolithic migration out of Africa, as supported by archeology (Bar Yosef 1987). Lumping the epipaleolithic remains of the Nile Valley and even those from the Maghreb, into one group has little to support it..."
- Keita, Studies and Comments on Ancient Egyptian Biological Relationships, 1993.
"Over 100 human skeletons of Late Paleolithic age are known from Egypt and adjacent Sudan. Physically, they are all classified as Homo Sapiens. They are grouped with the Mechtoids of the Maghreb, but details of their teeth indicate that they are a separate population, with many similarities to groups in sub-Saharan Africa."
- Encyclopedia of Prehistory - Volume 1: Africa Published in conjunction with the Human Relations Area Files (Encyclopedia of Prehistory) (Hardcover) by Peter N. Peregrine (Editor), Melvin Ember (Editor)Publisher: Springer; 1 edition (January 2001) p.117
|
|
|
|
Post by Dawn2Earth on Dec 21, 2010 19:56:46 GMT -5
Ancient Egypt Anatomy of a Civilisation(Paperback) by Barry Kemp (Author) Publisher: Routledge; 2 edition (December 12, 2005)
p.54
"sample populations available from northern Egypt from before the 1st Dynasty (Merimda, Maadi and Wadi Digla) turn out to be significantly different from sample populations from early Palestine and Byblos [...] If there was a south-north cline of variation along the Nile Valley it did not, from this limited evidence, continue smoothly
on into Palestine."
Side note: Here might lay the problem in getting evidence for inhabitants during that era in the low lying North:
Atlas of Ancient Egypt (Cultural Atlas of), John Baines; Hardcover page 59
"In the Paleolithic period [before 5000 B.C.] the Delta and Nile Valley were virtually uninhabitable. The annual flood [inundation] of the river Nile would have placed all areas of the Nile Valley under water for three months of each year, and at other times it was covered with thick vegetation that provided habitation for a variety of wild animals. In the north much of the low lying Delta was converted with papyrus swamps. At this time people lived on the desert spurs and hunted prolific game. As the climate became drier and the vegitation of the Nile Valley gradually changed , they were able to move down into the valley once the inudation receded . Here during the Neolithic period[c. 5000-4000 B.C.] they began to cultivate the land ,gorwing grain and learning to domesticate animals."
|
|
|
|
Post by Dawn2Earth on Dec 23, 2010 20:11:48 GMT -5
These three unrelated papers seem to be pointing in the same direction:
Who WeretheAncient Egyptians? Dental Affinities Among Neolithic Through Postdynastic Peoples
Joel D. Irish et. al. 2006
"Did Egyptians in the second half of the dynastic period become biologically distinct from those in the first?
Ideally, more dynastic samples than those from Abydos, Thebes, Qurneh, Tarkhan, Saqqara, Lisht, and Gizashouldbe compared to address such abroad question. Yet excluding the Lisht and perhaps Saqqara outliers, it appears that overall dental homogeneity among these samples would argue against such a possibility (Table4; Figs. 2,3,5). Specifically, an inspection of MMD values reveals no evidence of increasing phenetic distance between samples from the first and second halves of this almost 3,000-year-long period. For example, phenetic distances between First-Second Dynasty Abydos and samples from Fourth Dynasty Saqqara (MMD¼0.050), 11- 12thDynastyThebes (0.000), 12thDynastyLisht (0.072), 19thþDynastyQurneh (0.053), and 26th-30thDynasty Giza (0.027) do not exhibita directional increase through time. Moreover, there is no conspicuous correlation between MMD and geographic distances within and between Upper and Lower Egypt. A similar pattern is evident when comparing First Dynasty Tarkhan to these same five Old Kingdom through Late Dynastic samples. All display moderate frequencies of the nine influential traits identified by CA, and a largely concordant occurrence of, and trends across, the remaining traits (Table 2). Thus, despite increasing foreign influence after the Second Intermediate Period, not only did Egyptian culture remain intact (Lloyd, 2000a), but the people themselves, as represented by the dental samples, appear biologically constant as well. These findings coincide with those of Brace et al. (1993, p. 1), who stated that the Egyptians were ''largely unaffected by either invasions or migrations,'' and do not support suggestions of increased diversity due to infiltration of outside physical elements.
Did Egyptians of the Ptolemaic and Roman periods differ significantly from their dynastic antecedents?
Again, more postdynastic samples would prove useful in answering this broad question. Moreover, any foreign genetic influence on the indigenous populace likely diminished relative to the distance upriver. However, as it stands, the lone Greek Egyptian (GEG) sample from Lower Egypt significantly differs from all but the small Roman-period Kharga sample (Table 4). In fact, it was shown to be a major outlier that is divergent from all others (Figs. 2,3,5). The Greek Egyptians exhibit the lowest frequencies of UM1 cusp 5, three-rooted UM2, five-cusped LM2, and two-rooted LM2, along with a high incidence of UM3 absence, among others (Table 2). This trait combination is reminiscent of that in Europeans and western Asians (Turner, 1985a; Turner and Markowitz, 1990; Roler, 1992; Lipschultz, 1996; Irish, 1998a). Thus, if the present heterogeneous sample is at all representative of peoples during Ptolemaic times, it may suggest some measure of foreign admixture, at least in Lower Egypt near Saqqara and Manfalut. Another possibility is that the sample consists of actual Greeks. Although their total number was probably low (Peacock, 2000), Greek administrators and others were present in Lower Egypt. Future comparisons to actual Greek specimens will help verify this possibility."
"Egyptian continuity extends across time (as evidenced by affinities among the three predynastic, five of seven dynastic, and two or perhaps three Roman period samples) and space (as indicated by the mostly random distribution of points denoting Upper and Lower Egyptians)."
- Irish et. al. 2006
Facts gleaned from the study
* Continuity throughout the Dynastic era and across Kemet.
* The Greek period sample a major outlier only resembling the Roman period one.
So any major demic events have to occur prior to, late in, and after Dynastic times. Joel and the gang go on to write:
"Still, the patterning illustrated by the MDS and CA diagrams is of interest, and will receive attention in future studies comparing Egyptians to samples from elsewhere in northeast Africa, greater North Africa, sub- Saharan Africa, and the western Mediterranean area."
- Irish 2006
Another study takes a look at a different feature and reaches similar conclusions:
Zakrzewski, Sonia R. (2003) Variation in Ancient Egyptian Stature and Body Proportions. American Journal of Physical Anthropology 121(3): 219-229.
"The nature of the body plan was also investigated by comparing the intermembral, brachial, and crural indices for these samples with values obtained from the literature. No significant differences were found in either index through time for either sex. The raw values in Table 6 suggest that Egyptians had the “super-Negroid” body plan described by Robins (1983). The values for the brachial and crural indices show that the distal segments of each limb are longer relative to the proximal segments than in many “African” populations (data from Aiello and Dean, 1990). This pattern is supported by Figure 7 (a plot of population mean femoral and tibial lengths; data from Ruff, 1994), which indicates that the Egyptians generally have tropical body plans. Of the Egyptian samples, only the Badarian and Early Dynastic period populations have shorter tibiae than predicted from femoral length. Despite these differences, all samples lie relatively clustered together as compared to the other populations."
- Zakrzewski et. al. 2003
Comparisons with folks further down the Nile (which has one major source in Ethiopia and the other clear past the equator) ..
"Badarian occupies a position closest to the Teita, Gaboon, Nubian, and Nagada series by centroid values and territorial maps. The Nagada and the Kerma series are so similar that they are barely distinguishable in the territorial maps; they subsume the first dynasty series in Abydos… The Badarian crania have a modal metric phenotype that is clearly “southern”; most classify into the Kerma (Nubian), Gaboon, and Kenyan groups… No Badarian cranium in any analysis classified into the European series"
- Keita (1990)
|
|
|
|
Post by Dawn2Earth on Dec 23, 2010 20:16:07 GMT -5
Why "race" has fallen out of scientific use as far as mainstream biology is concerned: We tested the accuracy and effectiveness of Fordisc 2.0 using twelve cranial measurements from a homogeneous population from the X-Group period of Sudanese Nubia (350CE-550CE). When the Fordisc program classified the adult X-Group crania, only 51 (57.3%) of 89 individuals were classified within groups from Africa. Others were [/b]placed in such diverse groups as Polynesian (11.24%), European (7.86%), Japanese (4.49%), Native American (3.37%), Peruvian (3.36%), Australian (1.12), Tasmanian (1.12%), and Melanesian (1.12%).[/b] The implications of these findings suggest that classifying populations, whether by geography or by "race", is not morphologically or biologically accurate because of the wide variation even in homogeneous populations."[/i] - J. Edwards, A. Leathers, et al The idea was to seek to explain human variation through a given number of races and there mixing. Problems occur when there is no agreement on the number because East Indians have traits that can't be explained interms of black and white Westerners, who don't represent the whole Earth's human genome. Then you have three races only to find that Oceanians are the only folks left with a certain facial trait, or that the "pure" races you thought must exist due to having traits set at extremes appear or turn out to be "mixed" in other ways. Eventually the racial picture becomes so complex or the number so high your not far from extended families, making the whole concept of divisions arbitrary. You can see epicanthic eye folds ("slanted eyelids") in the Sahara, in South Africa, Nigeria, and black America -- they may not be as common, but that doesn't make everyone with such a trait the cousin of Jackie Chan. Overall analysis of the human genome has revealed for instance that for Europe, Asia, and Africa the vast majority of genetic variation is within them not between them -- only a small means exists outside of Africa that's not covered within infact. Not to mention genetic studies where Africa and even East Asia show up more as actual breeding populations than does say Europe which looks like a mixture between those two (clickable link to an image of the diagram, while Oceania (Pacific Islanders like in Fiji and the Andamanese) who live in the tropics, are relly black and are the spitting image of stereotypical africoid traits are *the* population out of all others that is most genetically unique from Africa. This is simply helped to Africa's vast genetic diversity, and other areas of the globe being less diverse the further away they are whilst also being basically a subset of Andamanese diversity. In the past there were scientists could triflently use idea of human races - before genetics revealed for taxonomists that subdivisions of the species HSS don't exist - as proxies withwhich to steal authenticity of other populations. |
|
|
|
Post by Dawn2Earth on Dec 26, 2010 13:59:31 GMT -5
Prime example, and one relevant specifically to the topic at hand:
TABLE 4. Intra-limb bone length indices
in US and Egyptian samples
********************* Crural indexA__ | ___Brachial indexB
******************* Males_ \ _Females | __Males___ \ _Females_
**************** Mean - SE \ Mean - SE | Mean - SE__ \ Mean - SE
****** U.S. Whites: 81.9 - 0.4 \ 82.0 - 0.4 | 74.3 - 0.4__ \ 73.5 - 0.5
****** U.S. Blacks: 83.7 - 0.4 \ 83.8 - 0.5 | 77.1 - 0.5_ \ 76.5 - 0.5
Ancient Egyptians: 83.6c - 0.2 \ 82.8 - 0.3 | 77.9c - 0.5 \ 77.5c - 0.6
- courtesy Raxter et. al. 2007
In different ways, overall Ancient Kemetians and U.S. Blacks measure further than one another from U.S. whites. None of these populations are a totem by which to measure purity however. But then neither is any population. Say we have a group of say Asians, as source population from which another group we'll call neo-Asians fully derive, and they diverge even further from collective non-Asians than had their source meaning either reduced phenetic variation held in common with non-Asians. This would appear to be a "purer race" than the population that spawned them and in this hypothetical scenario still by that time retained all the genetic variation they have and more. This is all why methods have to be revised and fine tuned; it's evolution and adaptation. We now in studies of physical remains use tactics like looking for traits that are not subject to positive or negative natural selection (i.e. evolution) and measure these closely to see if we see any change with any variables, like in the Zakrzewski et. al. 2007 study.
By the way -- brachial indices (intra-limb, as opposed to limb-trunk) place them well within the tropical African range -- but i imagine it's very hot in deserts, like a giant frying pan.
On a different note: did kemetians vary by class? Did royals, high officials, and farmers display any distinct patterns?
From the same study:
"There is no evidence for significant temporal or class-related variation among ancient Egyptians in linear body proportions. Thus, the new equations may be broadly applicable to Egyptian archaeological samples.
[...]
When plotted, proportions between workers and high officials also showed consistency and independent samples t-test results demonstrated no effect of social class on any of the proportions investigated (P [ 0.10)."
|
|
|
|
Post by truthcentric on Dec 28, 2010 12:09:49 GMT -5
By the way -- brachial indices (intra-limb, as opposed to limb-trunk) place them well within the tropical African range -- but i imagine it's very hot in deserts, like a giant frying pan. It does get hot during the daytime, but the thing about subtropical deserts is that they aren't hot 24/7. They cool down significantly during nighttime and the winter. That's why populations living in subtropical deserts for many millennia, such as the San people of southern Africa, have limb proportions intermediate between Europeans and other sub-Saharans.  |
|
|
|
Post by zarahan on Nov 10, 2011 16:59:03 GMT -5
The "African climate" incorporates diverse temperature, humidity, atmospheric pressure, wind, rainfall, atmospheric particle count and other meteorological elements in a wide range of environments -- from deserts, to high altitude snowy zones, to jungle, to savannah, to mixed woodlands, to higher altitude cloud forest, and all that is WITHIN the TROPICAL zone of Africa. Africa have the highest phenotypic diversity being where anatomically modern humans originated- that diversity is built-in to begin with, but side from this, the broad variation if climates and micro- climates WITHIN the tropical zone also means there will be variation based on environmental factors. No "race mix" is needed fundamentally to explain narrow noses for example. Narrow noses can occur in colder high altitude mountain zones, or colder high altitude cloud forest, or near cooler coastline zones or in hot deserts where noses adapt to the need to regulate temps of incoming air. While Africa has had migrants over centuries, and has had hybrids, like any other continent, no "incoming Caucasoids" are needed to fundamentally explain native variation already in place. Skin color likewise can vary.  --------------------------------------------------------- And just as tropical African environments are diverse,
so are tropical African peoples as credible scientists
note time and time again.QUOTES: Most phenotypic variation"Both methods for estimating regional diversity show sub-Saharan Africa to have the highest levels of phenotypic variation, consistent with many genetic studies." --- Relethford, John "Global Analysis of Regional Differences in Craniometric Diversity and Population Substructure". Human Biology - Volume 73, Number 5, October 2001, pp. 629-636) Most genetic variation"Africa contains tremendous cultural, linguistic and genetic diversity, and has more than 2,000 distinct ethnic groups and languages.. Studies using mitochondrial (mt)DNA and nuclear DNA markers consistently indicate that Africa is the most genetically diverse region of the world." ---Tishkoff SA, Williams SM., Genetic analysis of African populations: human evolution and complex disease. Nature Reviews Genetics. 2002 Aug (8):611-21.) Most skin color variation"Previous studies of genetic and craniometric traits have found higher levels of within-population diversity in sub-Saharan Africa compared to other geographic regions. This study examines regional differences in within-population diversity of human skin color. Published data on skin reflectance were collected for 98 male samples from eight geographic regions: sub-Saharan Africa, North Africa, Europe, West Asia, Southwest Asia, South Asia, Australasia, and the New World. Regional differences in local within-population diversity were examined using two measures of variability: the sample variance and the sample coefficient of variation. For both measures, the average level of within-population diversity is higher in sub-Saharan Africa than in other geographic regions. This difference persists even after adjusting for a correlation between within-population diversity and distance from the equator. Though affected by natural selection, skin color variation shows the same pattern of higher African diversity as found with other traits." -- Relethford JH.(2000). Human skin color diversity is highest in sub-Saharan African populations. Hum Biol. 2000 Oct;72(5):773-80.) |
|
|
|
Post by Dawn2Earth on Jan 14, 2013 4:30:39 GMT -5
By the way -- brachial indices (intra-limb, as opposed to limb-trunk) place them well within the tropical African range -- but i imagine it's very hot in deserts, like a giant frying pan. It does get hot during the daytime, but the thing about subtropical deserts is that they aren't hot 24/7. They cool down significantly during nighttime and the winter. That's why populations living in subtropical deserts for many millennia, such as the San people of southern Africa, have limb proportions intermediate between Europeans and other sub-Saharans. Very good point. I recognize that graph, it's from Stringer & Gamble 1993: crural index = Tibia/Femur length
modern peoples 79% in Lapps
86% in Black African groups
Group crural Mean annual temp C
index
average Neanderthal 79% ?
Modern peoples
Lapps 79% .25
modern Inuit 81.5% 4
Belgium 82.5% 10
S.African white 83.2% 8.5
Yugoslav 83.75% 8.4
American white 82.6% 9.8
Kalahari Bushman 83.4% 18
New MexicoIndian 84.6% 14
S.African black 86.4% 17
Arizona Indian 85.5% 18
Melanesian 84.8% 23
Pygmy 85.1% 24.2
Egyptian 84.9% 26.1
American Black 85.25% 26- Stringer and Gamble, 1993, p. 92 They get their Egyptian sample from Trinkhaus '81, the purpose of the study to compare Neanderthal to modern man. Here's what kind of Egyptians Trinkhaus used (he and his Egyptians are cited in the following citation):  - Keita 1993 |
|
|
|
Post by Dawn2Earth on Jan 15, 2013 12:58:21 GMT -5
Does anyone know about the cranial studies discussed at Egyptsearch where they found the cranial and or dental traits of groups changed during the Neolithic and switch to farming and different dietary habits?
Particularly one on the Nubians?
Or either, those studies existed but the one on the Nubians I'm looking for is the one where they cranially more resemble West Africans than later Nile Valley inhabitants. Thanks.
|
|
|
|
Post by clouddesignc7 on Sept 6, 2016 13:35:29 GMT -5
It does get hot during the daytime, but the thing about subtropical deserts is that they aren't hot 24/7. They cool down significantly during nighttime and the winter. That's why populations living in subtropical deserts for many millennia, such as the San people of southern Africa, have limb proportions intermediate between Europeans and other sub-Saharans. Very good point. I recognize that graph, it's from Stringer & Gamble 1993: crural index = Tibia/Femur length
modern peoples 79% in Lapps
86% in Black African groups
Group crural Mean annual temp C
index
average Neanderthal 79% ?
Modern peoples
Lapps 79% .25
modern Inuit 81.5% 4
Belgium 82.5% 10
S.African white 83.2% 8.5
Yugoslav 83.75% 8.4
American white 82.6% 9.8
Kalahari Bushman 83.4% 18
New MexicoIndian 84.6% 14
S.African black 86.4% 17
Arizona Indian 85.5% 18
Melanesian 84.8% 23
Pygmy 85.1% 24.2
Egyptian 84.9% 26.1
American Black 85.25% 26- Stringer and Gamble, 1993, p. 92 They get their Egyptian sample from Trinkhaus '81, the purpose of the study to compare Neanderthal to modern man. Here's what kind of Egyptians Trinkhaus used (he and his Egyptians are cited in the following citation): - Keita 1993
reposting incase the picture doesn't show:
Limb Ratio Studies
Limb ratio studies are of interest because of limb ratios' general relationship to climate as per Allen's rule. Mammals (including Homo sapiens sapiens) tend to have shorter distal members of the extremities in colder climates; this is viewed as being adaptive. Hence the shin (tibia)/thigh (femur) index in Europeans would on average be expected to differ from an equatorial population. Indeed, this is one line of evidence used to support the idea that at least some, if not most, Upper Paleolithic (anatomically modern) "Europeans" were immigrants from warmer areas (Trinkhaus 1981). Of course variation is expected in any region or population.
Trinkhaus (1981) provides upper and lower extremity distal/proximal member ratios for numerous populations, including a predynastic Egyptian and Mediterranean European series. The predynastic Egyptian values plotted near tropical Africans, not Mediterranean European.
- Keita 1993
|
|
|
|
Post by zarahan on Sept 28, 2016 18:34:54 GMT -5
Very good point. I recognize that graph, it's from Stringer & Gamble 1993: crural index = Tibia/Femur length
modern peoples 79% in Lapps
86% in Black African groups
Group crural Mean annual temp C
index
average Neanderthal 79% ?
Modern peoples
Lapps 79% .25
modern Inuit 81.5% 4
Belgium 82.5% 10
S.African white 83.2% 8.5
Yugoslav 83.75% 8.4
American white 82.6% 9.8
Kalahari Bushman 83.4% 18
New MexicoIndian 84.6% 14
S.African black 86.4% 17
Arizona Indian 85.5% 18
Melanesian 84.8% 23
Pygmy 85.1% 24.2
Egyptian 84.9% 26.1
American Black 85.25% 26- Stringer and Gamble, 1993, p. 92 They get their Egyptian sample from Trinkhaus '81, the purpose of the study to compare Neanderthal to modern man. Here's what kind of Egyptians Trinkhaus used (he and his Egyptians are cited in the following citation): - Keita 1993
reposting incase the picture doesn't show:
Limb Ratio Studies
Limb ratio studies are of interest because of limb ratios' general relationship to climate as per Allen's rule. Mammals (including Homo sapiens sapiens) tend to have shorter distal members of the extremities in colder climates; this is viewed as being adaptive. Hence the shin (tibia)/thigh (femur) index in Europeans would on average be expected to differ from an equatorial population. Indeed, this is one line of evidence used to support the idea that at least some, if not most, Upper Paleolithic (anatomically modern) "Europeans" were immigrants from warmer areas (Trinkhaus 1981). Of course variation is expected in any region or population.
Trinkhaus (1981) provides upper and lower extremity distal/proximal member ratios for numerous populations, including a predynastic Egyptian and Mediterranean European series. The predynastic Egyptian values plotted near tropical Africans, not Mediterranean European.
- Keita 1993
Good points. Some additional comments: 1) Almost 15-17% of AE is itself in the tropical zone. And that zone has fluctuated further north. 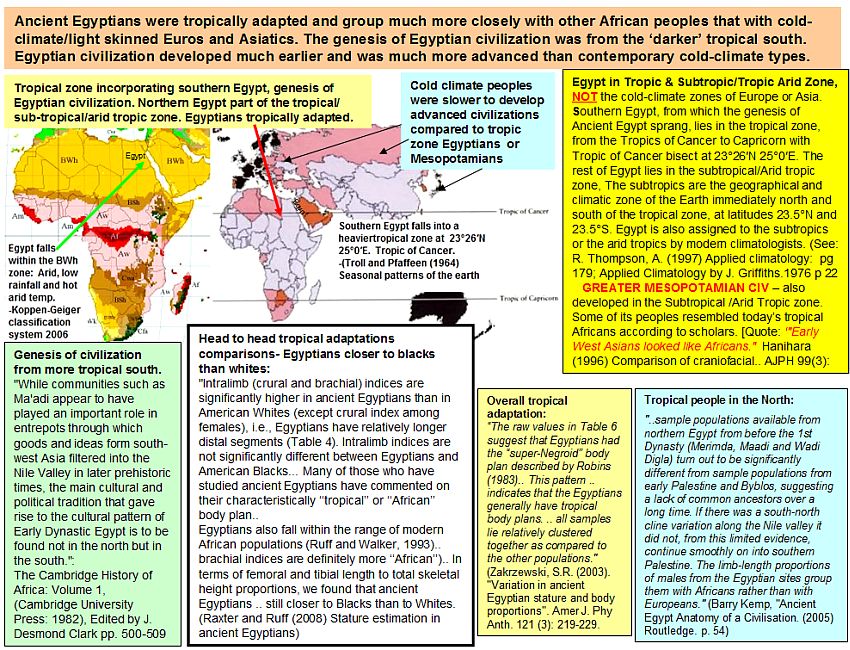 2) ^^African Americans are not a sub-tropical population- they are ancestrally a tropical one. 3) The San have intermediate indices in their cooler zone but they are all the same "SubSaharan" Africans, hard as it is for some people to believe. 4) While Africans show tropical adaptations they have also ranges in such things as crural indices that overlap European populations- and can be less than Europeans- an unsuprising pattern given the greater phenotypic diversity of Africans.  |
|
|
|
Post by clouddesignc7 on Oct 1, 2016 11:32:06 GMT -5
Prime example, and one relevant specifically to the topic at hand: TABLE 4. Intra-limb bone length indices
in US and Egyptian samples
********************* Crural indexA__ | ___Brachial indexB
******************* Males_ \ _Females | __Males___ \ _Females_
**************** Mean - SE \ Mean - SE | Mean - SE__ \ Mean - SE
****** U.S. Whites: 81.9 - 0.4 \ 82.0 - 0.4 | 74.3 - 0.4__ \ 73.5 - 0.5
****** U.S. Blacks: 83.7 - 0.4 \ 83.8 - 0.5 | 77.1 - 0.5_ \ 76.5 - 0.5
Ancient Egyptians: 83.6c - 0.2 \ 82.8 - 0.3 | 77.9c - 0.5 \ 77.5c - 0.6- courtesy Raxter et. al. 2007 By the way -- brachial indices (intra-limb, as opposed to limb-trunk) place them well within the tropical African range -- but i imagine it's very hot in deserts, like a giant frying pan. On a different note: did kemetians vary by class? Did royals, high officials, and farmers display any distinct patterns?From the same study: "There is no evidence for significant temporal or class-related variation among ancient Egyptians in linear body proportions. Thus, the new equations may be broadly applicable to Egyptian archaeological samples.
[...]
When plotted, proportions between workers and high officials also showed consistency and independent samples t-test results demonstrated no effect of social class on any of the proportions investigated (P [ 0.10)."
At least 95 out of 100 of their samples were from Lower Egypt too, right off the edge of the Delta:
******* Time period/location ************|* Male * Female
Predynastic (c. 3150--3050 BCE)/Keneh *****|*** 1 ***** 1
Predynastic (c. 3150--3050 BCE)/Masaeed ***|*** 1 ***** 0 Old Kingdom (c. 2687-2191 BCE)/Giza(b) ***|*** 50 **** 33 Old Kingdom (c. 2687-2191 BCE)/Giza(c) ***|**** 6 ***** 0 Middle Kingdom (c. 2061-1665 BCE)/Lisht ***|**** 2 ***** 2 New Kingdom (c. 1569-1081 BCE)/Lisht *****|**** 1 ***** 0 Late Period (c. 724-333 BCE)/Lisht *********|**** 0 ***** 1 Coptic (c. 337-641 CE)/Luxor **************|**** 2 ***** 0

|
|
|
|
Post by clouddesignc7 on Oct 5, 2016 11:56:38 GMT -5
F. X. Ricaut M. Waelkens
Cranial Discrete Traits in a Byzantine Population and Eastern Mediterranean Population Movements
Human Biology - Volume 80, Number 5, October 2008, pp. 535-564
Abstract: "Since the beginning of the Holocene, the Anatolian region has been a crossroads for populations and civilizations from Europe, Asia, and the Near to Middle East, with increasing interactions since the Bronze Age. In this context, we examine cranial discrete traits from a Byzantine population from southwest Turkey, excavated at the archeological site of Sagalassos; the site displays human occupation since the 12th millennium b.p. To investigate the biological history of this population, we analyzed the frequency distribution of 17 cranial discrete traits from Sagalassos and 27 Eurasian and African populations. Ward’s clustering procedure and multidimensional scaling analyses of the standardized mean measure of divergence (MMDst), based on trait frequencies, were used to represent the biological affinity between populations. Our results, considered within a large interpretive framework that takes into account the idea that populations are dynamic entities affected by various influences through time and space, revealed different strata of the Sagalassos biological history. Indeed, beyond an expected biological affinity of the Sagalassos population with eastern Mediterranean populations, we also detected affinities with sub-Saharan and northern and central European populations. We hypothesize that these affinity patterns in the Sagalassos biological package are the traces of the major migratory events that affected southwest Anatolia over the last millennia, as suggested from biological, archeological, and historical data."
"Keeping in mind these three elements, if we consider the affinity of the Sagalassos population with the sub-Saharan populations from Gabon and Somalia, a recent direct contact between these populations and regions probably can be excluded because they are seperated by significant geographic distances. However, indirect contacts through geographically intermediary populations carrying "sub-Saharan" biological features in the late Pleistocene-Holocene period are discussion points."
"From the Mesolithic to the early Neolithic period different lines of evidence support an out-of-Africa Mesolithic migration to the Levant by northeastern African groups that had biological affinities with sub-Saharan populations. From a genetic point of view, several recent genetic studies have shown that sub-Saharan genetic lineages (affiliated with the Y-chromosome PN2 clade; Underhill et al. 2004) have spread through Egypt into the Near East, the Mediterranean area, and, for some lineages, as far north as Turkey(E3b-M35 Y lineage; Cinnioglu et al. 2004; Luis et al. 2004), probably during several dispersal episodes since the Mesolithic (Cinnioglu et al. 2004; King et al. 2008; Lucotte and Mercier 2003; Luis et al. 2004; Quintanna-Murci et al. 1999; Semino et al. 2004; Underhill et al. 2001). This finding is in agreement with morphological data that suggest that populations with sub-Saharan morphological elements were present in northeastern Africa, from the Paleolithic to at least the early Holocene, and diffused northward to the Levant and Anatolia beginning in the Mesolithic. Indeed, the rare and incomplete 33,000-year-old Nazlet Khater specimen (Pinhasi and Semal 2000), the Wadi Kubbaniya skeleton from the late Paleolithic site in the upper Nile Valley (Wendorf et al. 1986), the Qarunian (Faiyum) early Neolithic crania (Henneberg et al. 1989; Midant-Reynes 2000), and the Nabta specimen from the Neolithic Nabta Playa site in the western desert of Egypt (Henneberg et al. 1980)-show, with regard to the great African biological diversity, similarities with some of the sub-Saharan middle Paleolithic and modern sub-Saharan specimens. This affinity pattern between ancient Egyptians and sub-Saharans has also been noticed by several other investigators (Angel 1972; Berry and Berry 1967, 1972; Keita 1995) and has been recently reinforced by the study of Brace et al. (2005), which clearly shows that the cranial morphology of prehistoric and recent northeast African populations is linked to sub-Saharan populations (Niger-Congo populations). These results support the hypothesis that some of the Paleolithic-early Holocene populations from northeast Africa were probably descendents of sub-Saharan ancestral populations."
"A late Pleistocene-early Holocene northward migration (from Africa to the Levant and to Anatolia) of these populations has been hypothesized from skeletal data (Angel 1972, 1973; Brace 2005) and from archaeological data, as indicated by the probable Nile Valley origin of the "Mesolithic" (epi-Paleolithic) Mushabi culture found in the Levant (Bar Yosef 1987). This migration finds some support in the presence in Mediterranean populations (Sicily, Greece, southern Turkey, etc.; Patrinos et al.; Schiliro et al. 1990) of the Benin sickle cell haplotype. This haplotype originated in West Africa and is probably associated with the spread of malaria to southern Europe through an eastern Mediterranean route (Salares et al. 2004)following the expansion of both human and mosquito populations brought about by the advent of the Neolithic transition (Hume et al 2003; Joy et al. 2003; Rich et al 1998). This northward migration of northeastern African populations carrying sub-Saharan biological elements is concordant with the morphological homogeneity of the Natufian populations (Bocquentin 2003), which present morphological affinity with sub-Saharan populations (Angel 1972; Brace et al. 2005). In addition, the Neolithic revolution was assumed to arise in the late Pleistocene Natufians and subsequently spread into Anatolia and Europe (Bar-Yosef 2002), and the first Anatolian farmers, Neolithic to Bronze Age Mediterraneans and to some degree other Neolithic-Bronze Age Europeans, show morphological affinities with the Natufians (and indirectly with sub-Saharan populations; Angel 1972; Brace et al 2005), in concordance with a process of demic diffusion accompanying the extension of the Neolithic revolution (Cavalli-Sforza et al. 1994)."
"Following the numerous interactions among eastern Mediterranean and Levantine populations and regions, caused by the introduction of agriculture from the Levant into Anatolia and southeastern Europe, there was, beginning in the Bronze Age, a period of increasing interactions in the eastern Mediterranean, mainly during the Greek, Roman, and Islamic periods. These interactions resulted in the development of trading networks, military campaigns, and settler colonization. Major changes took place during this period, which may have accentuated or diluted the sub-Saharan components of earlier Anatolian populations. The second option seems more likely, because even though the population from Sagalassos territory was interacting with northeastern African and Levantine populations [trade relationships with Egypt (Arndt et al. 2003), involvement of thousands of mercanries from Pisidia (Sagalassos region) in the war around 300 B.C. between the Ptolemaic kingdom (centered in Egypt) and the Seleucid kingdom (Syria/Mesopotamia/Anatolia), etc.], the major cultural and population interactions involving the Anatolian populations since the Bronze Age occured with the Mediterranean populations form southeastern Europe, as suggested from historical and genetic data."
"Consequently, one may hypothesize as the most parsimonious explanation that sub-Saharan biological elements were introduced into the Anatolian populations after the Neolithic spread and have been preserved since this time, at least until the 11th-13th century A.D., in the population living in the Sagalassos territory of southwestern Anatolia. This scenario implies that the affinity between Sagalassos and the two sub-Saharan populations (Gabon and Somalia) is more likely due to the sharing of a common ancestor and that the major changes and increasing interactions in the eastern Mediterranean beginning in the Bronze Age did not erase some of the sub-Saharan elements carried by Anatolian populations, as shown by genetic data and the morphologivcal features of our southwestern Anatolian sample."
"In this context it is likely that Bronze Age events may have facilitated the southward diffusion of populations carrying northern and central European biological elements and may have contributed to some degree of admixture between northern and central Europeans and Anatolians, and on a larger scale, between northeastern Mediterraneans and Anatolians. Even if we do not know which populations were involved, historical and archaeological data suggest, for instance, the 2nd millenium B.C. Minoan and later Mycenaean occupation of Anatolian coast, the arrival in Anatolia in the early 1st millennium B.C. of the Phrygians coming from Thrace, and later the arrival of settlers from Macedonia in Pisidia and in the Sagalassos territory (under Seleucid rule). The coming of the Dorians from Northern Greece and central Europe (the Dorians are claimed to be one of the main groups at the origin of the ancient Greeks) may have also brought northern and central European biological elements into southern populations. Indeed, the Dorians may have migrated southward to the Peloponnese, across the southern Aegean and Create, and later reached Asia Minor."
|
|
|
|
Post by clouddesignc7 on Oct 8, 2016 14:49:03 GMT -5
Reposting, without the rest of it:
F. X. Ricaut M. Waelkens
Cranial Discrete Traits in a Byzantine Population and Eastern Mediterranean Population Movements
Human Biology - Volume 80, Number 5, October 2008, pp. 535-564
"From the Mesolithic to the early Neolithic period different lines of evidence support an out-of-Africa Mesolithic migration to the Levant by northeastern African groups that had biological affinities with sub-Saharan populations. From a genetic point of view, several recent genetic studies have shown that sub-Saharan genetic lineages (affiliated with the Y-chromosome PN2 clade; Underhill et al. 2004) have spread through Egypt into the Near East, the Mediterranean area, and, for some lineages, as far north as Turkey(E3b-M35 Y lineage; Cinnioglu et al. 2004; Luis et al. 2004), probably during several dispersal episodes since the Mesolithic (Cinnioglu et al. 2004; King et al. 2008; Lucotte and Mercier 2003; Luis et al. 2004; Quintanna-Murci et al. 1999; Semino et al. 2004; Underhill et al. 2001). This finding is in agreement with morphological data that suggest that populations with sub-Saharan morphological elements were present in northeastern Africa, from the Paleolithic to at least the early Holocene, and diffused northward to the Levant and Anatolia beginning in the Mesolithic. Indeed, the rare and incomplete 33,000-year-old Nazlet Khater specimen (Pinhasi and Semal 2000), the Wadi Kubbaniya skeleton from the late Paleolithic site in the upper Nile Valley (Wendorf et al. 1986), the Qarunian (Faiyum) early Neolithic crania (Henneberg et al. 1989; Midant-Reynes 2000), and the Nabta specimen from the Neolithic Nabta Playa site in the western desert of Egypt (Henneberg et al. 1980)-show, with regard to the great African biological diversity, similarities with some of the sub-Saharan middle Paleolithic and modern sub-Saharan specimens. This affinity pattern between ancient Egyptians and sub-Saharans has also been noticed by several other investigators (Angel 1972; Berry and Berry 1967, 1972; Keita 1995) and has been recently reinforced by the study of Brace et al. (2005), which clearly shows that the cranial morphology of prehistoric and recent northeast African populations is linked to sub-Saharan populations (Niger-Congo populations). These results support the hypothesis that some of the Paleolithic-early Holocene populations from northeast Africa were probably descendents of sub-Saharan ancestral populations."[/b]
although the rest of it adds a backdrop especially for those of us familiar with the subject (of African genes found in Europe and the Mid-East).
|
|





