 DATA RECAP:
DATA RECAP:
2010 Berber mtDNA study finds Berber roots foundational in Africa - frigi 2010Ancient Local Evolution of African mtDNA Haplogroups in Tunisian Berber Populations
Frigi et al.
Human Biology
August 2010 (82:4)
Discussion
In this study we attempted to better elucidate the ancient African genetic
background in the northwest African area, particularly in Tunisia. To this aim, we
focused our study on Berber populations that are considered representative of the
ancient North African populations that probably derived from Neolithic Capsians.
During historic times, Berbers experienced a long and complicated history with
many invasions, conquests, and migrations by Phoenicians, Romans, Vandals,
Byzantines, Arabs, Bedouins, Spanish, Turks, Andalusians, sub-Saharans (communities
settled in Jerba and Gabes in the 16th–19th centuries), and French (Brett
and Fentress 1996). During these invasions, Berbers were forced back to the mountains
and to certain villages in southern Tunisia (Fadhlaoui-Zid et al. 2004).
At present, they are restricted to some isolates in the south who maintain the Berber
language and to some populations in the north who lack an origin language.
Many genetic studies on Tunisian Berber populations demonstrate the heterogeneity
of Berbers with respect to European and sub-Saharan African contributions
and the mosaic structure of Tunisian Berber populations with an absence of ethnic,
linguistic, and geographic effects (Cherni et al. 2010).
In the present work, mtDNA data show a diversified distribution of African
haplogroups. However, a question remains concerning the date of the sub-Saharan
African inputs. Our results demonstrate an ancient local evolution in Tunisia of
some African haplogroups (L2a, L3*, and L3b). The most ancient haplogroup is
L3*, which would have been introduced from eastern sub-Saharan populations to
North Africa about 20,000 years ago. The Siwa oasis sample studied by Coudray
et al. (2009) contains sub-Saharan haplogroups L0a1, L3i, L4*, and L4b2, which
are different from our Tunisian samples, in agreement with the heterogeneity of
Berbers already shown in Tunisia.
Stevanovitch et al. (2004) suggested that the Gurna population in Egypt has
conserved the trace of an ancestral genetic structure from an ancestral East African
population characterized by a high haplogroup M1 frequency. This haplogroup is
also present in three Berber populations (Kesra, Matmata, and Sned) with variable
frequencies. In each of these populations, haplogroup L3* is also present.
The association of both eastern African haplogroups in the Berber populations is
a strong argument in favor of eastern African gene flow in Berbers. Other genetic
and archaeological studies confirmed the crucial idea that an ancient population in
East Africa constituted the basis of the ancestors of all African Upper Paleolithic
populations—and their subsequent present-day descendants (Bengtson 2008; Keita
2004; Relethford 2000; Zakrzewski 2003, 2007).

Moreover, Berber languages spoken exclusively by North African populations
belong to the Afro-Asiatic language. Diakonoff (1998) showed an exclusively
African origin (Diakonoff, 1981, 1988) for the family. He explicitly described
proto-Afro-Asiatic vocabulary as consistent with non-food-producing vocabulary
and linked it to pre-Neolithic cultures in the Levant and in Africa south of Egypt.
Moreover, Ehret. (2003) suggested that early Afro-Asiatic languages were spread
by Mesolithic foragers from Africa into the Levant. On the contrary, Diamond
and Bellwood (2003) suggested that food production and the Afro-Asiatic
language family were brought simultaneously from the Near East to Africa by demic
diffusion—in other words, by a migration of food-producing peoples. The evidence
presented by Wetterstrom (1993) does not support this latter suggestion,
however, and indicates that early African farmers in the Fayum initially incorporated
Near Eastern domesticates into an existing indigenous foraging strategy and
only over time developed a dependence on horticulture.
In conclusion, the crucial linguistic finding is that the three deepest clades
of the Afro-Asiatic family are localized in Eritrea and Ethiopia. All the other languages
of the family outside that region belong to subclades of just one of those
deep clades. This kind of cladistic distribution is a basic criterion of the
genetic argument for the genetic lineage origins well understood by geneticists. It applies
to linguistic history as well.
Our results also point to a less ancient western African gene flow to Tunisia
involving haplogroups L2a and L3b. Thus the sub-Saharan contribution to northern
Africa starting from the east would have taken place before the Neolithic. The
western African contribution to North Africa should have occurred before the Sahara’s
formation (15,000 years BP). It seems likely that an expansion would have
taken place in the Sahel zone starting about the time of a gradual climatic return
to wetter conditions, when the Senegal River cut through the dunes (Burke et al.
1971). For subhaplogroup L2a1 (data not shown) we found some haplotypes that
the Tunisian Berbers shared with Mauritanians and western sub-Saharan populations
speaking a Niger-Congo language (studied by Salas et al. 2002).
This suggests that the people who brought these markers to the Berber populations most
likely came from West African populations that spoke languages belonging to the
Niger-Congo family when the Sahara became drier. However, this contribution
of West African haplotypes and of other haplotypes, such as those belonging to
haplogroup L1b1, could have been introduced to North Africa more recently.
Indeed, this West African contribution was difficult to date, because few
haplotypes belonging to western African haplogroups have been observed, most
of them being divergent. This result can be interpreted in different ways. Ancient
western African mtDNA contributions could have disappeared from North Africa
as a result of recent flows, or the situation observed now could be the result of a
strong drift effect on ancient western African lineages, particularly those belonging
to haplogroups L2a and L3b. A strong Iberian gene flow may have contributed
to the decrease in African haplogroups. Indeed, most of the older hypotheses
about North African population settlement used to suppose an Iberian or an eastern
origin. The dates for subhaplogroups H1 and H3 (13,000 and 10,000 years,
respectively) in Iberian and North African populations allow for this possibility.
Kefi et al.’s (2005) data on ancient DNA could be viewed as being in agreement
with such a presence in North Africa in ancient times (about 15,000–6,000 years
ago) and with the fact that the North African populations are considered by most
scholars as having their closest relations with European and Asian populations
(Cherni et al. 2008; Ennafaa et al. 2009; Kefi et al. 2005; Rando et al. 1998).
However, considering the general understanding nowadays that human settlement of
the rest of the world emerged from eastern northern Africa less than 50,000 years
ago, a better explanation of these haplogroups might be that their frequencies reflect
the original modern human population of these parts of Africa as much as or
more than intrusions from outside the continent. The ways that gene frequencies
may increase or decrease based on adaptive selection, gene flow, and/or social
processes is under study and would benefit from the results of studies on autosomal
and Y-chromosome markers.
Since the end of the extreme Saharan desiccation, lasting from before
25,000 years ago up to about 15,000 years ago, the Sahara has had post- and pre-
Holocene cyclical climatic changes (Street and Grove 1976), and corresponding increases
and decreases in population are probable. Wetter phases with better habitats
perhaps allowed for increased colonization and gene and cultural exchange.
Desiccation would have encouraged the emigration and segmentation of populations,
with resultant genetic consequences secondary to drift producing more
variation. During the last glacial period, the Sahara was even bigger than it is
today, extending south beyond its current boundaries (Ehret 2002). About 13,000
years ago, large parts of the Sahara were as dry as the desert is now (White and
Mattingly 2006). The end of the glacial period brought more rain to the Sahara,
especially from about 8500 to 6000 BC (Fezzan Project 2006). By around 3400
BC, the monsoon retreated south to approximately where it is today, leading to the
gradual desertification of the region (Kröpelin 2008). Thus the Sahara, through its
cyclical environmental changes, might be seen as a microevolutionary “processor”
and/or “pump” of African people that “ejected” groups to the circum-Saharan
regions in times of increasing aridity.
Indeed, it must be noted that the high frequencies of cDe, P, and V antigens
and low frequencies of FY antigens in some Berber-speaking groups (Chamla
1980; Mourant et al. 1976) indicate affinities with tropical Africans. These data
may indicate recent or ancient gene flow from sub-Saharan Africa, a common immediate
pre-Holocene ancestral group, or chance resemblance.
Our findings are in accordance with other studies on Y-chromosome markers
that have shown that the predominant Y-chromosome lineage in Berber communities
is the subhaplogroup E1b1b1b (E-M81), which emerged in Africa, is
specific to North African populations, and is almost absent in Europe, except in
Iberia (Spain and Portugal) and Sicily. Molecular studies on the Y chromosome in
North Africa are interpreted as indicating that the southern part of Africa, namely,
the Horn/East Africa, was a major source of population in the Nile Valley and
northwest Africa after the Last Glacial Maximum, with some migration into the
Near East and southern Europe (Bosch et al. 2001; Underhill et al. 2001).
Hence, contrary to the suggestion that mtDNA haplogroups were introduced
mostly from Iberia, it seems that Y-chromosome markers have an eastern
African origin with an ancient local evolution in North Africa. These observations
are in agreement with the proposal that the ancient communities ancestral
in language to more recent Berber communities absorbed a lot of females from
the existing pre-Holocene populations. This would indicate that the North African
populations arose from admixture rather than from local evolution, leading
to an intermediate genetic structure between eastern sub-Saharan Africans and
Eurasians. Rock paintings in North Africa that show people of different phenotypes
living together are a strong argument for our hypothesis (Hachid 1982,
1992, 1998).
In conclusion, our findings parallel the more recent findings of both archaeology
and linguistics on the prehistory of Africa. The present study suggests that
sub-Saharan contributions to North Africa have experienced several complex population
processes after the occupation of the region by anatomically modern humans.
Our results reveal that Berber speakers have a foundational biogeographic root in
Africa and that deep African lineages have continued to evolve in supra-Saharan Africa.
-------------------------------------------------------------------------------------
Originally posted by Ish Gebor:
Berbers of Libya, Ghadamès.


Nafusa, of Jebel Nafusa in Tripolitania.


 www.britishmuseum.org/research/research_projects/all_current_projects/sudan/berber-abidiya_project.aspxwww.britishmuseum.org/research/research_projects/all_current_projects/berber-abidiya_project/the_berber-adiya_region.aspx
www.britishmuseum.org/research/research_projects/all_current_projects/sudan/berber-abidiya_project.aspxwww.britishmuseum.org/research/research_projects/all_current_projects/berber-abidiya_project/the_berber-adiya_region.aspx-------------------------







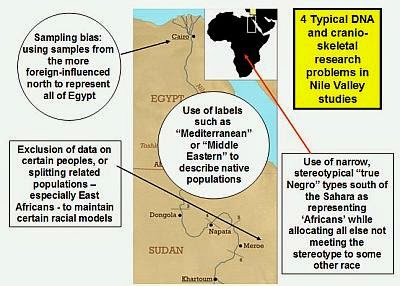
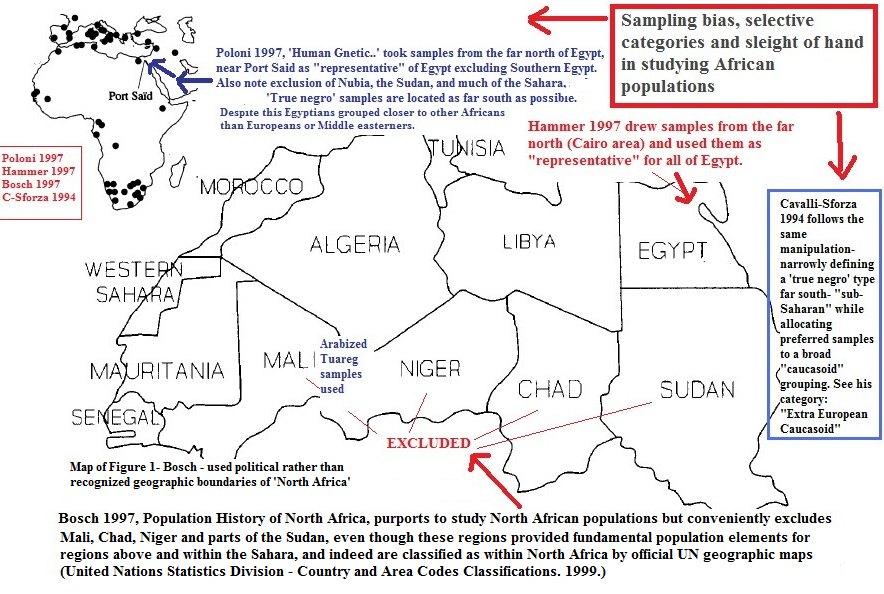

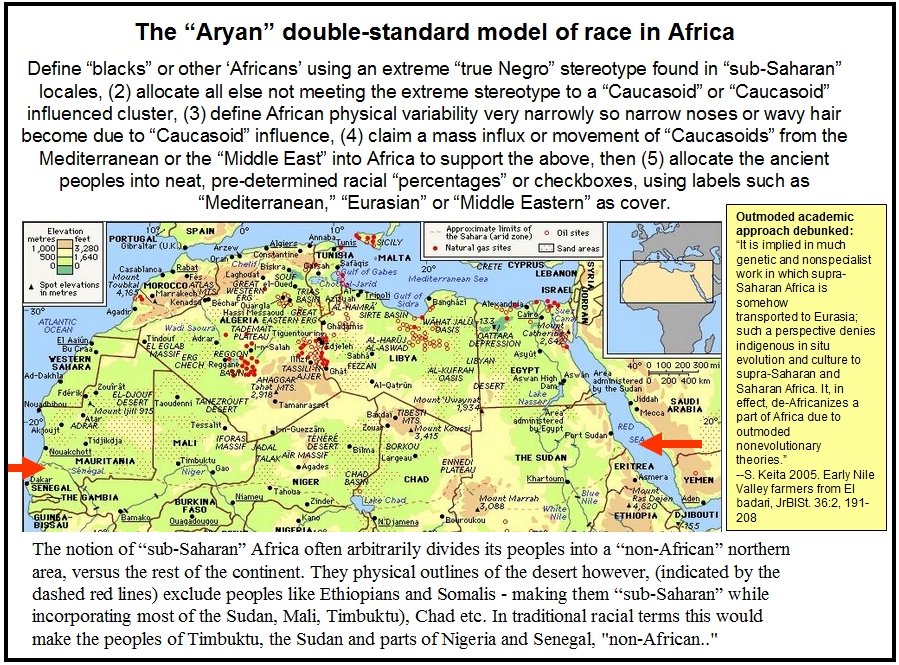
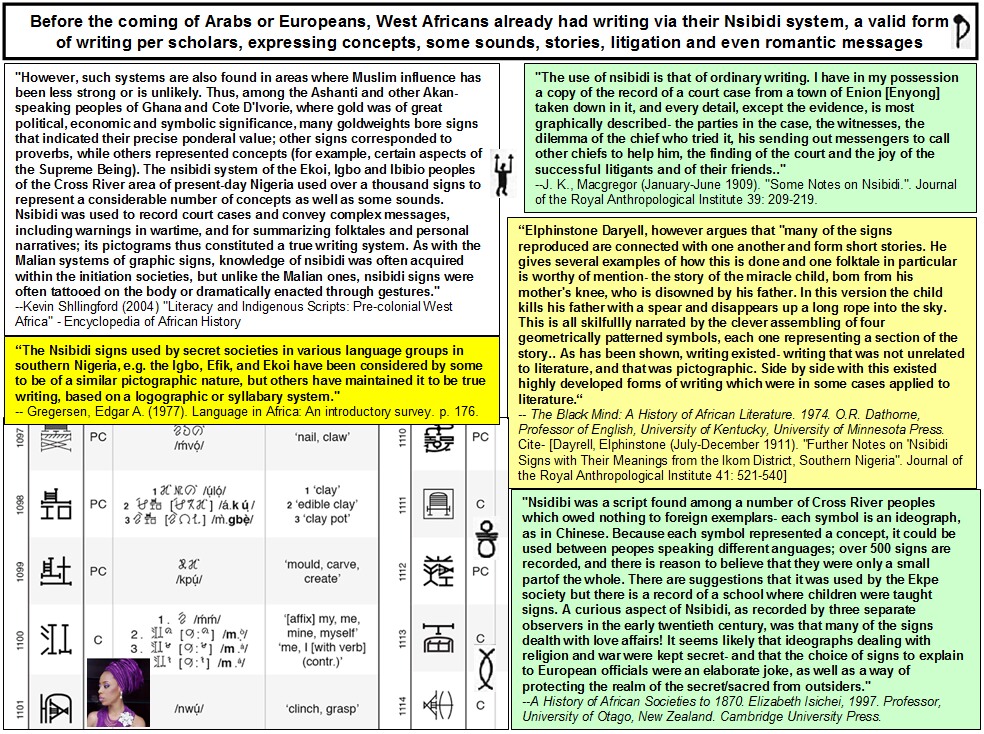








 [/QB]
[/QB]












